|
Discussion
 The synonymy of the treated taxa is generally straightforward, but Rissoa
corilea "d'Orbigny" G. B. Sowerby II, 1876, here considered a synonym of
Stosicia aberrans, warrants some review. The type illustration
[fig. 4]
is accompanied by a scale line 5.8 mm. long, and Cuba is given as the
habitat. Tryon (1887; pp. 363-364) considered Sowerby's name a simple
misspelling of d'Orbigny's Rissoa caribaea and used d'Orbigny's figure
(1842[?]; t. 11, bis fig. 32) to illustrate the two taxa. However, that
action misidentifies Rissoa corilea. Although Sowerby's illustration and
text feature the anterior canal, there is no such character in the type figure
of Rissoa caribaea d'Orbigny, who gives (1842; pp. 21-22)
the size of his
shell as "2 millim." and mentions "deux carènes... bandes brun... Bouche ovale"
which clearly relate to the species presently known as
Simulamerelina caribaea
(d'Orbigny, 1842), which is not even a congener [but see De Jong and Coomans,
1988 and Redfern, 2001 for differing interpretations at the species level].
Rosenberg (2003 et a priori) was the first to point out this peculiar
synonymy. The synonymy of the treated taxa is generally straightforward, but Rissoa
corilea "d'Orbigny" G. B. Sowerby II, 1876, here considered a synonym of
Stosicia aberrans, warrants some review. The type illustration
[fig. 4]
is accompanied by a scale line 5.8 mm. long, and Cuba is given as the
habitat. Tryon (1887; pp. 363-364) considered Sowerby's name a simple
misspelling of d'Orbigny's Rissoa caribaea and used d'Orbigny's figure
(1842[?]; t. 11, bis fig. 32) to illustrate the two taxa. However, that
action misidentifies Rissoa corilea. Although Sowerby's illustration and
text feature the anterior canal, there is no such character in the type figure
of Rissoa caribaea d'Orbigny, who gives (1842; pp. 21-22)
the size of his
shell as "2 millim." and mentions "deux carènes... bandes brun... Bouche ovale"
which clearly relate to the species presently known as
Simulamerelina caribaea
(d'Orbigny, 1842), which is not even a congener [but see De Jong and Coomans,
1988 and Redfern, 2001 for differing interpretations at the species level].
Rosenberg (2003 et a priori) was the first to point out this peculiar
synonymy.
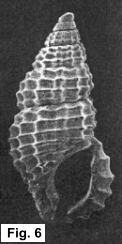
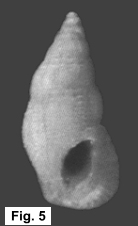
The taxonomic and zoogeographic relationships between these two Stosicia
species and among all the congeners invite speculation as to the evolutionary
scenario of their origin and deployment. Clearly S. aberrans and S.
houbricki are quite closely related; the illustrations of these and other
Stosicia in Sleurs (1996) attest to this propinquity. It therefore seems
likely that the two share a common ancestor. I looked high and low for any
fossil evidence of the presence of Stosicia (in any of its conceivable
nomenclatorial iterations) in the western Atlantic but could find none. Thus,
it also seems likely that the ancestor was probably an immigrant from either the
Eastern Atlantic or the Indo-West-Pacific in post-Miocene times, likely not long
ago at all. On analysis of protoconchs, if the immigrant ancestor were closer to R. aberrans,
it would have planktotrophic larvae with the ability to disperse over large
oceanic expanses unlike the case with S. houbricki with its inferred
lecithotrophic (crawl-away) larvae. What little we know of the ancestral forms
in Stosicia (e.g., S. planaxoides Grateloup, 1838, a senior synonym
of the type species; see Sleurs) suggests that they had multispiral protoconchs
(planktonic larvae) like that of S. aberrans, yet many of the Recent taxa
from the Indo-Pacific (and S. houbricki) have paucispiral protoconchs,
which infer a lecithotrophic larval habit (crawl-away young). Which came first
in our evolutionary scenario?
If
we look at other groups in which sympatric pairs of closely-related marine
snails having utterly different protoconchs much as do our two species, a few
instances in the Caribbean fauna leap to mind in which the teleoconchs are
actually even more similar: Alvania auberiana (d’Orbigny, 1842) / A.
faberi De Jong and Coomans, 1988, Retilaskeya emersonii (C. B. Adams,
1839) / R. bicolor (C. B. Adams, 1845), and Iniforis turristhomae
(Holten, 1802) / I. casta (Hinds, 1845). In these cases of so-called
didymous species pairs, it is not entirely clear to me which is the ancestral
and which the derived condition, but, in a group taxonomically close to the
Stosicia, Verduin (1977) looked at European Rissoa species and
concluded that the lecithotrophic species was the descendant (apomorphic), and
the planktotrophic was ancestral (plesiomorphic). Similar conclusions were
reached in a wider taxonomic and/or paleontological context in Ficus
(Smith, 1945), Trophon (Bouchet and Warén, 1985), Nassariidae (Martinell
and Cuadras, 1977), Neogastropoda in the early Tertiary and Volutidae in the
Cenozoic to Recent (Hansen, 1983), Terebridae (Bouchet, 1981), and Turridae (Bouchet,
1990). To my knowledge, no instances of the opposite pattern of evolution has
been reported. In fact, Strathman (1978) concluded that plankotrophic
development was ancestral to the lecithotrophic condition among all marine
invertebrates with such life histories, and that the reversal of this process
was much less frequent, even unlikely. A relatively limited geographic
distribution of lecithotrophic species and their tendency to be less long-lived
(in a geological sense) than their planktotrophic congeners has been noted by
several workers including Jablonski (1982, 1986) and Hansen (1983).
Thus, taking into account the evidence above and considering the substantial
range S. aberrans and probable recent appearance of Stosicia in
western Atlantic waters, we may create a scenario of planktonic larval waifs
swimming into the Caribbean, aided by the Gulfstream from the eastern side of
the Atlantic (where its parent stock subsequently perished) or, more likely,
based on geochronology, invading America from the Indo-West Pacific and
traversing the straits that were to become the isthmus of Panama sometime in,
say, the late Pliocene epoch. It then dispersed widely and rapidly. Somewhat
later a mutant population, the new species S. houbricki, arose in the
northern Caribbean. That new species spread on a lesser scale, now occupying a
smaller geographic range with perhaps its most prodigious leap being across the
Gulf Stream to southeastern Florida. It probably has very similar habits except
for its reproductive strategy. Having crawl-away young may endow that new
species with a selective advantage which allows it to prosper yet not outcompete
the sympatric (?syntopic) S. aberrans, the plesiomorphic form borne as a
juvenile in the plankton and thus more akin to, quite possibly conspecific with,
the immigrant ancestor. Likely the same pattern of speciation accounts for the
deployment of the eight lecithotrophic species of Stosicia in the
Indo-Pacific, half of which have limited ranges [see:
Stosicia species]. However, four of these non-planktotrophic
species [see: Stosicia incisa (Laseron, 1956)] have rather wide geographic distribution possibly reflecting an earlier
origin in geologic time, a factor not unexpected in a basin where the genus is
known to have a fossil record dating to the Miocene.
Bibliography
Adams, C. B., 1850. Descriptions of supposed new
species of marine shells which inhabit Jamaica. Contributions to Conchology no. 7:109-123. April, p.113; description of Rissoa aberrans.
[not seen; but see Clench and Turner, 1950].
Bavay, A. 1922. Sables littoraux de la Mer des Antilles provenant des abords de
Colon et de Cuba. Bulletin du Muséum national d'Histoire naturelle, Paris, 28(6):423-428.
[p. 424-425; fig. 1; Colon, Caribbean Panama].
Bouchet, P., 1981. Evolution of larval development in eastern Atlantic
Terebridae. Neogene to Recent. Malacologia 2: 363-369.
Bouchet, P., 1990. Turrid genera and mode of development: the use
and abuse of protoconch morphology. Malacologia 32(1): 69-77.
Bouchet, P. and A. Warén, 1985. Revision of the northeast Atlantic bathyal
and abyssal Neogastropoda, excluding Turridae. Bolletino Malacologico
supplement 1: 121-296.
Clench, W. J., C. G. Aguayo, and R. D. Turner, 1947-1948. The West Indian marine
shells by Henry Krebs (a republication by W. J. Clench, C. G. Aguayo, and R. D.
Turner). Revista de la Sociedad Malacologica "Carlos de la Torre" 5(1):
23-40; 5(2): 59-80; 5(2) 91-116; 6(1): 11-43; 6(2):45-48.
Clench, W. J. and R. D. Turner, 1950. The western Atlantic marine mollusks
described by C. B. Adams. Occ. Pap. Moll. 1(15): 233-403 incl. pls.
29-49. June 26. [p. 249; pl. 33, fig 10; holotype MCZ 156426; Jamaica].
De Jong, K. M. and H. E. Coomans, 1988. Marine gastropods from Curaçao,
Aruba and Bonaire. E. J. Brill, Leiden, v + pp 1-261 incl. 47 pls.
Diaz, J. M. and M. Puyana H[egedus]., 1994. Moluscos del Caribe
Colombiano. Un catálogo ilustrado. Fundación Natura, Sta. Fe de Bogota,
Colombia, pp. 1-291 + 78 pls.
d'Orbigny, A. 1853 [1842]. Mollusques. In: Ramon de la Sagra ed.
Histoire Physique, Politique et Naturelle de l'île de Cuba 2: 1-112, pls.
10-21? Arthus Bertrand: Paris. [Stated date 1853]
Espinosa, J., R. Fernández-Garcés, and E. Rolán, 1994. Catálogo de los moluscos
marinos actuales de Cuba. Cuadernos de investigación biológica (Bilbao) 18:
85-154.
Espinosa, J. and J. Ortea, 2002. Description of four new species of the
family Rissoinidae (Mollusca: Gastropoda). Avicennia 15: 141-146. [p.
144; fig. 1C, two views; holotype and one paratype Instituto de Oceanología,
Havana; one paratype Museo de Ciencias Naturales de Tenerife, Canary Is.].
Hansen, T., 1983. Modes of larval development and modes of speciation in
early Tertiary neogastropods. Science 204: 501-502.
Houbrick., J. R. [R. S.], 1968. A survey of the littoral marine mollusks
of the Caribbean coast of Costa Rica. The Veliger 11(1): 4-23. July 1.
Jablonski, D., 1982. Evolutionary rates and modes in late Cretaceous
gastropods: role of larval ecology. Proceedings of the Third North American
Geological Convention 1: 257-262. [not seen]
Jablonski, D., 1986. Larval ecology and macroevolution in marine invertebrates.
Bulletin of Marine Science 39: 565-587. [not seen]
Krebs, H., 1864. The West Indian marine shells with some remarks.
W. Laubs Widow & Chr. Jorgensen, Nykøbing, Falster, Denmark pp. 1-137.
[not seen; but see Clench, Aguayo, and Turner, 1947-1948.]
Martinell, J. and C. Cuadras, 1977. Bioestadistica y analysis multivariable
aplicados a la comparacion de una poblacion actual y otra fosil, atribuidas a
Sphaeronassa mutabilis (Linné) aportacion a la sistematica del genero
Sphaeronassa Locard, 1886. Studia Geologica (Salamanca) 13:
89-103. [not seen]
Mørch, O. A. L., 1876. Synopsis molluscorum marinorum Indiarum Occidentalum
imprimus Insularum Danicarum. Malakozoologische Blätter 23: 87-143.
Nowell-Usticke, G. W., 1959. A checklist of the marine shells of St.
Croix, U. S. Virgin Islands, with random notations. Lane Press, Burlington,
VT, vi + pp. 1-90 + 4 pls.
Odé, H., 1986. Distribution and records of the marine Mollusca in the
northwest Gulf of Mexico (A continuing monograph). Texas Conchologist 23(1): 20-36. Oct.
[Family Rissoidae].
Olsson, A. A. and T. L. McGinty, 1958. Recent marine mollusks from the
Caribbean coast of Panama with the description of some new genera and species.
Bull. Amer. Paleont. 39(177): 1-58.
Pilsbry, H. A., 1939. Land Mollusca of North America (north of Mexico) vol. 1
part 1. Academy of Natural Sciences, Philadelphia. xvii + pp 1- 573 + ix.
Pointier, J. P. and D. Lamy, 1998. Guide des coquillages des Antilles.
PLB, Guadeloupe. pp 1-223 incl. numerous color text figures + map + 2 pp.
afterleaf (unpaginated). Sept.
Ponder, W. F., 1984. A review of the genera of the Rissoidae (Mollusca:
Mesogastropoda: Rissoacea). Records of the Australian Museum supplement
4: 1-221.
Radwin, G. E., 1969. A Recent molluscan fauna from the Caribbean coast of
southeastern Panama. San Diego Soc. Nat. Hist. Trans. 15(14): 227-236.
June 27.
Redfern, C., 2001. Bahamian seashells a thousand species from Abaco,
Bahamas. Bahamianseashells.com, Inc., Boca Raton, pp. 1-280 + ix + 120 pls.
Rios, E. C., 1994. Seashells of Brasil 2a edição. Fundação Cidade do Rio
Grande, Rio Grande do Sul, Brasil, pp. 1-368 + 113 pls.
Rosenberg, G., 2003 Malacolog version 3.2.3 Western Atlantic Gastropod
Database.
July 3.
Sleurs, W. J. M., 1989. A zoogeographical analysis of the Rissoinine fauna
of the Eastern Pacific with special reference to a comparison with the Caribbean
fauna and with a checklist of the Eastern Pacific Rissoininae Stimpson, 1865 (Mollusca:
Gastropoda). Ann. Soc. Roy. Zool. Belg. 19(2): 155-164 [not seen].
Sleurs, W.J.M., 1996. A revision of the recent species of the genus
Stosicia
(Gastropoda: Rissoidae). Academiae Analecta Brussels 58(1): 115-158
with 19 plate-like figures [pp. 142-144, pl. 14, figs. f-i; holotype and 8
paratypes USNM 775230; 6 paratypes USNM 757703 all from reef flat, Carrie Bow
Key, Belize]
Smith, B., 1945. Observations on gastropod
protoconchs. Paleontographica Americana 3(19): 1-48 [225-268] + 3
pls. Sept. 11.
Sowerby, G. B. II, 1876. Monograph of the genus Rissoa.
Conchologia Iconica 20. [sp. 24; pl. 3, fig. 24; description of Rissoa
corilea "d'Orbigny"].
Strathman, R., 1978. The evolution and loss of feeding larval stages of
marine invertebrates. Evolution 32: 894-906.
Tryon, G. W., 1887 Manual of conchology; structural and systematic 9:
1-488 + 71 plates. Academy of Natural Sciences, Philadelphia.
Turgeon, D. D. and W. G. Lyons, 1978. A tropical marine assemblage in the
northeast Gulf of Mexico. Bull. Amer. Malac. Union (1977): 88-89
abstract.
Verduin, A., 1977. On a remarkable dimorphism of the apices in many groups
of sympatric, closely-related marine gastropod species. Basteria 41: 91-95.
Vokes, H. E. and E. H. Vokes, 1983 [1984]. Distribution of shallow-water
marine Mollusca, Yucatan Peninsula, Mexico. Mesoamerican Ecological
Institute, Monograph 1, Middle American Research Institute, Publication 54:
i-viii + 1-183, incl. 50.
Warmke G. L. and R. T. Abbott, 1961. Caribbean Seashells.
Livingston Publishing Co., Narberth, PA, xx + pp. 1-348, incl. 44 pls.
Acknowledgements:
The author thanks Bill Frank for technical and editorial
assistance - in particular for his fine digital photography and other work on
the illustrations. Paul Callomon, John Chesler, Dr. Emilio García, William G.
Lyons, Bob Pace, Richard E.
Petit, Colin Redfern, Dr. Gary Rosenberg, John Slapcinsky, and Peggy Williams
deserve my thanks for provision of specimens, records, or images.
Epilogue:
A few years after the above report was posted,
more and better specimens of western Atlantic Stosicia came to
light, including some shells collected in Honduras by frequent web site
contributor, Emilio García. After close study of this additional
material, it became clear that the shells of S. houbricki and
S. fernandezgarcesi consistently differed and that Emilio's
shells were distinct from those two taxa and S. aberrans.
Appropriately, the fourth western Atlantic congener was dubbed
Stosicia garciai Rolán, Fernández-Garcés
and Lee, 2009.
Rolán E., R. Fernández-Garcés, and H.G. Lee, 2009. The genus Stosicia
in the Caribbean (Caenogastropoda, Rissoidae) with the description
of a new species. Basteria 73(1-3): 1-8. |
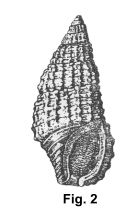
 In April, 1850
Charles Baker Adams (1814-1853) named Rissoa aberrans based on a
shell collected in Jamaica, possibly by himself, during a visit in the
winter of 1848-49, just five years before he died of yellow fever on his
third Caribbean field trip. It appears that Prof. Adams, writing from his
desk at Amherst College, selected the trivial name because of a
conchological character atypical for the genus in which he placed the
taxon, an anterior (siphonal) canal in the aperture. He wrote "This
species connects the genus with those Cerithia, in which the canal is
reduced to a notch." In fact this feature calls to mind Cerithium
lutosum Menke, 1828, which at first glance might be mistaken for a
giant specimen of Adams' species. A century later Clench and Turner
(1950) illustrated the holotype (and, for that matter, Adams' species) for
the first time
In April, 1850
Charles Baker Adams (1814-1853) named Rissoa aberrans based on a
shell collected in Jamaica, possibly by himself, during a visit in the
winter of 1848-49, just five years before he died of yellow fever on his
third Caribbean field trip. It appears that Prof. Adams, writing from his
desk at Amherst College, selected the trivial name because of a
conchological character atypical for the genus in which he placed the
taxon, an anterior (siphonal) canal in the aperture. He wrote "This
species connects the genus with those Cerithia, in which the canal is
reduced to a notch." In fact this feature calls to mind Cerithium
lutosum Menke, 1828, which at first glance might be mistaken for a
giant specimen of Adams' species. A century later Clench and Turner
(1950) illustrated the holotype (and, for that matter, Adams' species) for
the first time  Rissoa buccinalis
Grateloup, 1828) and the central Pacific, Stosicia Brusina, 1878.
He placed it with the genera Rissoina and Zebina in the
Rissoininae, a subfamily of the Rissoidae. Stocisia aberrans (C.
B. Adams, 1850) was said to be the only member of its genus surviving in
the Atlantic Ocean. Unfortunately he appears to have lacked intact
material as he strongly implied the species had a paucispiral protoconch (vide
infra).
Rissoa buccinalis
Grateloup, 1828) and the central Pacific, Stosicia Brusina, 1878.
He placed it with the genera Rissoina and Zebina in the
Rissoininae, a subfamily of the Rissoidae. Stocisia aberrans (C.
B. Adams, 1850) was said to be the only member of its genus surviving in
the Atlantic Ocean. Unfortunately he appears to have lacked intact
material as he strongly implied the species had a paucispiral protoconch (vide
infra).